
Abstract
Direct rhizogenesis from leaf explants and establishment of an in vitro stolon culture system and subsequent plant regeneration for Glycyrrhiza glabra have been described. MS liquid medium supplemented with 0.01 mg l−1 of NAA was most effective for stolon proliferation. Extensive proliferation of stolon and shoot regeneration was achieved on medium containing 3 % sucrose with 0.01 mg l−1 NAA. Stolons with nodes showing growth was transferred under light for plantlet regeneration in the same medium. This paper is the first report in G. glabra describing a complete regeneration procedure via in vitro stolon proliferation along with quantitative data for glycyrrhizin and genetic fidelity of plant regenerated in vitro there from. In vitro stolon proliferation described here would be an efficient way for regeneration of plants for functional genomics studies and better understanding of glycyrrhizin (GA) metabolism.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Glycyrrhiza (Fabaceae), commonly known as licorice, is an important source of flavonoids and terpenoids class of compounds, important among them is glycyrrhizin (GA). Roots and stolons of Glycyrrhiza plants (G. uralensis Fisch. and G. glabra L., Fabaceae) are highly valued commercial products used in oriental medicine (Anonymous 2005). In modern medicine it is considered an important crude drug for its various pharmacological activities (Jatav et al. 2011) including anti-diabetic, anti-inflammatory (Finney and Somers 1958), hepato-protective, anti-ulcer and anti-allergic (Park et al. 2004). Glycyrrhizin also has antiviral activity (Fiore et al. 2008) against various DNA and RNA viruses, including HIV (Ito et al. 1988) and severe acute respiratory syndrome (SARS)-associated corona virus (Cinatl et al. 2003). In recent studies it is shown to selectively block the tumor COX-2 pathway and suppress colon carcinogenesis in mice and humans (Zhang et al. 2009).
Glycyrrhizin, an oleanane type terpenoid saponin, and its derivative compounds are exclusively obtained from the dried roots and stolons of licorice. Chemical analyses have failed to detect them in the aerial part (Hayashi et al. 1988); the plants have been indiscriminately exploited to meet the high demand, resulting in desertification of the habitat. The plant is naturally found in areas restricted to Central and South China, Russia and Mediterranean regions. Under cultivation, the plant is rarely propagated by seeds due to its hard testa (Gupta et al. 1997).Vegetative propagation of the plant is annual and germination percentage is low, being highly under the influence of environmental conditions.
Developing in vitro regeneration methods of Glycyrrhiza glabra utilizing different explants viz, stem segments (Thengane et al. 1998) and axillary bud (Shah and Dalal 1980) to produce multiple shoots (Henry et al. 1991), shoot regeneration from callus (Wongwicha et al. 2008) shoots differentiation and clonal propagation (Mousa et al. 2006) has been the subject of several studies. Recent study by Mehrotra et al. (2012) has demonstrated the potential of encapsulated in vitro-grown axillary micro-shoots of Glycyrrhiza glabra in long-term storage and year round availability of germplasm for commercial cultivation. Hairy root induction using Agrobacterium rhizogenes have been recently developed in G. glabra (Li et al. 2000; Shirazi et al. 2012) showing the feasibility to cultivate these transformed roots in air lift bioreactors (Mehrotra et al. 2008). Tissue culture techniques have raised expectation regarding possibility of their application for propagation and understanding of glycyrrhizin biosynthesis. Tissue culture method to propagate G. glabra on commercial scale has also been reported by Shah and Dalal (1980). Recent studies regarding glycyrrhizin production by in vitro cultures have shown contrasting and conflicting results. Most of the attempts to detect glycyrrhizin from cultured tissues of G. glabra have been unsuccessful (Hayashi et al. 1988; Saito et al. 1990; Ayabe et al. 1990; Toivonen and Rosenqvist 1995). Callus and cell suspension cultures of selected G. glabra clones have shown production of glycyrrhizin, detected using TLC-tentative analysis (Nagat et al. 2007). Similarly, Wongwicha et al. (2008) reported glycyrrhizin production in callus cultures of G. uralensis (36.52 mg g−1), G. glabra (14.39 mg g−1), and G. inflata (16.14 mg g−1).
Although development of in vitro regeneration procedures utilizing various explants in G. glabra have been the subject of many studies, most of these reports lacked information on the molecular stability and biochemistry of the plant regenerated using tissue culture procedures. Inter-simple sequence repeats (ISSR) was chosen as molecular marker in the present study because this technique relies on the high polymorphism and can detect low differentiation levels having more stringent primer annealing conditions than RAPDs leading to higher accuracy and reproducibility (Zietkiewicz et al. 1994).
The present study focuses on utilization of root culture, stolon proliferation for high-throughput plant regeneration and analyze glycyrrhizin and related terpenoids accumulation in root cultures and in vitro-proliferated stolons. Additionally, molecular analysis of regenerated plants using ISSR markers have also been undertaken to assess genetic uniformity.
Materials and methods
Plant material
A selected clone of G. glabra plants collected from agricultural farms of Himachal Pradesh Agricultural University, Solan, located in the north Himalayan region (30.92°N and 77.15°E) of Indian subcontinent was utilized for the present study.
Direct rhizogenesis from foliar explants
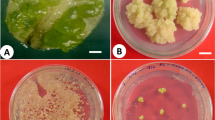
Fully expanded young leaves of the G. glabra plant were collected and washed under tap water. Whole leaf was surface sterilized with 0.1 % HgCl2 for 5 min. The leaves were further rinsed with sterile water five times and inoculated on culture medium. Plantlets were regenerated in vitro using shooting and rooting mediums. The leaves of the in vitro raised plants were used for studying the prospect of direct root initiation and plant regeneration. Young, soft leaves measuring 0.5–1 cm2 were obtained from 5-week-old in vitro raised plantlets along with petiole under aseptic conditions and cultured on MS (Murashige and Skoog 1962) medium containing 3 % sucrose and supplemented with cascade of growth regulators (IBA, Kinetin, IAA and 2,4-D) as shown in Table 1. The pH of the medium was adjusted to 5.8 and autoclaved at 121 °C under pressure of 1.2 kg cm−1 for 15 min. First observation was recorded after 14 days of culturing and subsequent observations were made every week thereon. Primary cultures showing rhizogenesis (Fig. 1a, b) were transferred on to stolen proliferation liquid MS medium (3 % sucrose, 0.01 mg l−1 NAA), incubated in dark and grown at 28° ± 2 °C on a rotary shaker at 100 rpm.

In vitro stolon organogenesis and plant regeneration from cultured stolons in Glychyrriza glabra. a Adventitious root induction from leaf explants; b root cultures and stolon proliferation from cultured roots in MS medium containing 3 % sucrose, 0.01 mg l−1 NAA; c, d stolon segments and shoot differentiation with plantlet regeneration; e, f regenerated plantlets after acclimatization (3 months); g ISSR profile of ten randomly selected regenerated plants from roots; h LC–MS chromatogram showing peaks of glycyrrhizin analysis of cultured stolons
De novo shoot proliferation from cultured stolons
The stolon cultures grown in dark at 28 ± 2 °C under shaking conditions were transferred to light under a 16-h photoperiod provided by cool white fluorescent tubes (Phillips, India) having a light intensity of 40 μmol m−2 s−1 (Fig. 1c, d)
Transfer of plant to soil
Plantlets with well developed shoot and root systems were transferred to the green house for hardening. Roots of plantlets regenerated in vitro were washed thoroughly to remove agar and planted into plastic pots containing sterilized sand–soil mixture (2:1). They were kept covered for the first 2 weeks and watered twice a week in the first week using ½ MS stock solution followed by sterile water in the subsequent week on the alternative days (Fig. 1e, f)
Molecular and biochemical analysis
Molecular analysis
The plants regenerated via stolons were selected randomly for assessing genetic stability using five ISSR markers. Young fresh leaves of the ten selected in vitro regenerated plants, hardened under glass house conditions, were utilized for molecular analysis. High-quality DNA was extracted following CTAB method (Doyle and Doyle 1987) with slight modifications. The quality and quantity of the DNA was ascertained by spectrophotometer (Nanodrop, Thermo scientific) and agarose gel electrophoresis. PCR reactions were carried out in a 10× reaction buffer with 1.5 mM MgCl2, 1 mM dNTP, 10 pmol of primer, 25 ng of genomic DNA for 20 μl reaction volume with 1 Unit of Taq DNA polymerase. Amplification was performed in a thermal cycler (Eppendorf, Vapo protect), with one cycle of initial denaturation at 95 °C for 5 min, followed by 34 cycles at 94 °C for 30 s, annealing at their respective temperature (40–45 °C) for 2 min and extension at 72 °C for 1 min followed by final extension at 72 °C for 8 min. PCR products were separated on 1.5 % agarose gel containing 1× TAE and stained with ethidium bromide (0.5 μg ml−1). A 100 bp DNA ladder (Fermentas) was used as a standard molecular weight marker. Banding pattern was photographed and stored as digital picture in gel documentation system (Syngene, USA). Each experiment was repeated thrice (Fig. 1g).
Biochemical analysis
Chemicals and reagents
ACS certified HPLC-grade solvents supplied by JT Baker, India, were used in the analysis. Pure glycyrrhizin (GA) was purchased from Sigma, India. Water was purified using Milli-Q-plus filter systems (Millipore, Bedford, MA, USA).
Ultrasound-assisted extraction of glycyrrhizin
Green and brown stolons and root tuft were weighed and crushed using mortar, extracted for 10 min using 2 ml of 50 % ethanol under ultra-sonication (Elma, Germany), for 30 min at 45 MHz, at room temperature, twice. The extract was filtered (Millipore filter 0.45 μm) concentrated on a rotary-evaporator (Buchi, Switzerland) and 2 ml of solvent was added to the sample before analysis.
Glycyrrhizin detection and quantification
For the identification and quantification of glycyrrhizin present in in vitro raised stolons, a UPLC-qTOF-MS system (Synapt, Waters, USA, equipped with MassLynx acquisition software, version 4.1) was used. The system consisted of: column, C-18 (50 mm × 2.1 mm) with particle size 1.7 μm (Acquity, BEH) having mobile phase of acetonitrile and acetic acid (2 %) in water (75:25, v/v), at a flow rate of 200 μl min−1. The injection volume was 1 μl. Separation of the analyte was achieved at retention time of 1.17 min. Column and sample temperature were 30 and 10 °C, respectively. Infusion experiments were performed using an in-built syringe pump. A UPLC–qTOF-MS with ESI interface was used for MS/MS analysis, with following parameters: capillary voltage, 2.7 kV for positive mode; source temperature, 100 °C; desolvation temperature, 200 °C; cone gas flow, 50 l h−1 and desolvation gas flow, 550 l h−1. The multiple reaction monitoring (MRM) mode was used to monitor the transition of glycyrrhizin mz −1 845.1873 [M+Na], 867.1644 [M+K] 823.2040 [M+H] (Fig. 1h).
Results and discussion
Roots could be initiated on MS medium enriched with three different growth hormones (IBA, NAA and Kn) displaying well-grown, white roots after 2 weeks of culturing of leaf explants. Roots mostly induced at the proximal end of the whole leaf cultured on medium containing combination of Kn (1.0 mg l−1) with NAA or IBA (1.0 mg l−1) and IAA (1.0 mg l−1) alone (Fig. 1a, b). Addition of cytokinin or BAP along with IAA or NAA resulted in synergistic influence on evoking response on the leaf segments. The root culture proliferates very rapidly on transferring roots in liquid medium with 0.01 mg l−1 NAA, with the passage of time roots transferred to liquid medium assumed a mat like appearance within a period of 3 weeks. Repeated sub-culturing demonstrated autonomy of root meristem which grew vigorously as a whitish cottony mass under dark incubation. Among the five different PGR combinations—(1) NAA (1.0 mg l−1) + Kn (1.0 mg l−1); (2) IBA (1.0 mg l−1) + Kn (1.0 mg l−1); (3) NAA (1.0 mg l−1); (4) IAA (1.0 mg l−1) and (5) 2,4-D (1.0 mg l−1)—root development was most vigorous in MS medium supplemented with IAA or IBA with kinetin (Table 1). With the passage of time elongated roots slowly got transformed into thick and stout pale white or light brown stolons in medium under dark conditions. Several shoot primordial could be observed on stolons within 3 weeks on MS medium containing 3 % sucrose with 1.0 mg l−1 NAA. Medium supplemented with NAA (Fig. 1c) was found most suitable for shoot differentiation. Root differentiation and whole plantlet regeneration was observed after 2 subculture passages within 3 weeks when kept under light (Fig. 1c, d). Cultured stolons retained characteristic color similar to field grown stolons. Shoot bud and its differentiation in well developed shoots occurred mostly on middle portion of in vitro stolon which indicated that certain degree of maturity of stolon is required prior to shoot differentiation. On transferring, segments of cultured stolons to MS media treatment containing NAA (0.01 mg l−1) resulted in >8 shoots from each stolon segment (3 cm). The shoots obtained were found to root efficiently and resulted in complete plantlets. These were further maintained on the same medium under illumination. Plantlets were successfully acclimatized to soil in pots with 70 % survival, plants grew normal and vigorously under green house conditions (Fig. 1e, f; Table 2).The present described procedure is being reported for the first time for G. glabra where root cultures established in vitro could be effectively utilized to retrieve high-frequency plant regeneration via stolon proliferation. The present results provide protocol for complete in vitro regeneration utilizing excised in vitro regenerated roots leading to stolon proliferation and subsequent in vitro regeneration in G. glabra. Using present procedure 20–25 plants/stolon could be produced within 3 months. The root cultures are advantageous over other explants in terms of their easy manipulation and high regenerative potential (Franklin et al. 2004). However, such report in other species of Glycyrrhiza (G. uralensis) is available (Kojoma et al. 2010). The present described method holds promise in being simple, reproducible and high throughput for mass cloning of ‘elite’ genotypes of G. glabra. The application of developed procedure could be in providing large number of G. glabra plants in comparison to earlier published reports (Shah and Dalal 1980; Henry et al. 1991; Mousa et al. 2006).
Regenerated plantlets derived through stolon proliferation were subjected to molecular analysis in order to evaluate their genetic uniformity and/or detect likely existing variations among in vitro-derived plantlets. Ten ISSR primers were used for evaluating ISSR fingerprinting successfully validated true-to- type of in vitro raised regenerates (Fig. 1g). As such the described method could be used for mass cloning with minimum possibility of any in vitro-induced variability.
Glycyrrhizin production in three stages of in vitro raised stolons (young, mature and root tuft) confirmed the presence of glycyrrhizin in cultivated stolons using sensitive LC–MS analytical technique. However, quantitative differences in the glycyrrhizin content were observed in all the three stages. It was interesting to note that green immature stolon produced minimum glycyrrhizin (4.0 μg g−1) while maximum content (4.7 μg g−1) was detected in mature brown stolon in the present study (Table 3). Previous reports of Kojoma et al. (2010) detected the glycyrrhizin production in cultured stolons of G. uralensis support our present results. However, earlier published reports of glycyrrhizin production in tissue cultures of G. glabra were conflicting. Yoo and Kim (1976) reported glycyrrhizin production by callus of G. glabra. Later Hayashi et al. (1988), Ayabe et al. (1990) and Saito et al. (1990) failed to detect any glycyrrhizin in G. glabra.
Conclusion
In conclusion, the present described strategies were found to be most effective in induction of stolons which proliferated and extended high-throughput regeneration procedure in Glycyrrhiza glabra to obtain ideal true-to-type regenerants. Accumulation of glycyrrhizin in root cultures of G glabra and in vitro stolon system extends uniform tissue cultures which could be utilized for functional genomics and understanding glycyrrhizin biosynthetic studies. This would broaden available knowledge regarding the biosynthetic pathway and mechanism responsible for glycyrrhizin metabolism and its regulation. The mechanism and the biosynthetic enzymes responsible for its over production is still an open area for research.
Author contribution
R.A.V. is responsible for conceptualizing the idea. A.A. designed the experiments. S.G., P.P. were responsible for establishment of tissue culture experiment, chemical analysis was done by A.P.G. and M.K.V. A.A. and S.G. are also responsible for compiling the data in the form of MS.
Abbreviations
- 2,4-D:
-
2,4-Dichlorophenoxy acetic acid
- BA:
-
6-Benzyladenine
- HPLC:
-
High performance liquid chromatography
- IAA:
-
Indole-3-acetic acid
- IBA:
-
Indole-3-butyric acid
- KN:
-
Kinetin
- MS:
-
Murashige and Skoog medium
- NAA:
-
1-Naphthaleneacetic acid
- μM:
-
Micro mole
- ISSR:
-
Inter-simple sequence repeats
- LCMS:
-
Liquid chromatography–mass spectrometry
- GA:
-
Glycyrrhizin
- DNA:
-
Deoxyribonucleotide triphosphate
- RNA:
-
Ribonucleic acid
- SARS:
-
Severe acute respiratory syndrome
- COX-2:
-
Cyclo-oxygenase-2
- TLC:
-
Thin layer chromatography
- PCR:
-
Polymerase chain reaction
- CTAB:
-
Cetyl trimethylammonium bromide
- dNTP:
-
Di-nucleotide phosphate
- TAE:
-
Tris-acetate-ethylenediaminetetraacetic acid
References
Anonymous (2005) A dictionary of Indian raw materials and industrial products, first supplement series. In: The wealth of India. National Institute of Sciences Communication and Information Resources, CSIR, New Delhi, 3, pp 195–198
Ayabe S, Takano H, Fujita T, Furuya T, Hirota H, Takahashi T (1990) Triterpenoid biosynthesis in tissue cultures of Glycyrrhiza glabra var. glandulifera. Plant Cell Rep 9:181–184
Cinatl J, Morgenstern B, Bauer G, Chandra P, Rabenau H, Doerr HW (2003) Glycyrrhizin, an active component of liquorice roots, and replication of SARS-associated coronavirus. Lancet 361:2045–2046
Doyle JJ, Doyle JL (1987) A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem Bull 19:11–15
Finney RSH, Somers GF (1958) The antiinflammatory activity of glycyrrhetinic acid and derivatives. J Pharm Pharmacol 10:613–620
Fiore C, Eisenhut M, Krausse R, Ragazzi E, Pellati D, Armanini D, Bielenberg J (2008) Antiviral effects of Glycyrrhiza species. Phytother Res 22:141–148
Franklin G, Sheeba CJ, Laksmisita G (2004) Regeneration of eggplant (Solanum melongena L.) from root explants. In Vitro Cell Dev Biol Plant 40:188–191
Gupta V, Kak A, Singh BB (1997) Studies on seed germination and seedling vigour in liquorice (Glycyrrhiza glabra). JMAPS 2:412–413
Hayashi H, Fukui H, Tabata M (1988) Examination of triterpenoids produced by callus and cell suspension cultures of Glycyrrhiza glabra. Plant Cell Rep 7:508–511
Henry M, Edy MA, Desmarest P, Manoir DJ (1991) Glycyrrhiza glabra L. (Licorice): cell culture, regeneration, and the production of glycyrrhizin. In: Bajaj YPS (ed) Biotechnology in agriculture and forestry, vol 15, Medicinal and aromatic plants III. Springer, Berlin, pp 270–282
Ito M, Sato A, Hirabayashi K, Tanabe F, Shigeta S, Baba M, Clercq ED, Nakashima H, Yamamoto N (1988) Mechanism of inhibitory effect of glycyrrhizin on replication of human immunodeficiency virus (HIV). Antiviral Res 10:289–298
Jatav VS, Singh SK, Khatri P, Sharma AK (2011) Recent pharmacological trends of Glycyrrhiza glabra Linn. Unani Res 1:1–4
Kojoma M, Ohyama K, Seki H, Hiraoka Y, Asazu SN, Sawa S, Sekizaki H, Yoshida S, Muranaka T (2010) In vitro proliferation and triterpenoid characteristics of licorice (Glycyrrhiza uralensis Fischer, Leguminosae) stolons. Plant Biotechnol 27:59–66
Li W, Asad Y, Yoshikawa T (2000) Flavonoid constituents from Glycyrrhiza glabra hairy root cultures. Phytochemistry 55:447–456
Mehrotra S, Kukreja AK, Khanuja SPS, Mishra BN (2008) Genetic transformation studies and scale up of hairy root culture of Glycyrrhiza glabra in bioreactor. Electron J Biotechnol 11:1–7
Mehrotra S, Khwaja O, Kukreja AK, Rahman L (2012) ISSR and RAPD based evaluation of genetic stability of encapsulated micro shoots of Glycyrrhiza glabra following six months of storage. Mol Biotechnol 52:262–268
Mousa N, Siagura P, Wiryowidagdo S, Wagih ME (2006) Rapid clonal propagation of licorice (Glycyrrhiza glabra) by in vitro shoot culture. Sugar Tech 8:292–298
Nagat A, Mousa TR, Siaguru I, Wiryowidagdo S, Wagih ME (2007) Establishment of regenerative callus and cell suspension system of licorice (Glycyrrhiza glabra) for the production of the sweetener glycyrrhizin in vitro. Sugar Tech 9:72–82
Park HY, Park SH, Yoon HK, Han MJ, Kim DH (2004) Anti-allergic activity of 18β-glycyrrhetinic acid-3-O-beta-d glucuronide. Arch Pharm Res 27:57–60
Saito K, Kaneko H, Yamazaki M, Yoshida M, Murakoshi I (1990) Stable transfer and expression of chimeric genes in licorice (Glycyrrhiza uralensis) using a Ri plasmid binary vector. Plant Cell Rep 8:718–721
Shah RR, Dalal KC (1980) In vitro multiplication of Glycyrrhiza. Curr Sci 49:69–71
Shirazi Z, Piri K, Asl AM, Hasanloo T (2012) Glycyrrhizin and isoliquiritigenin production by hairy root culture of Glycyrrhiza glabra. J Med Plants Res 6:4640–4646
Thengane R, Kulkarni DK, Krishnamurthy KV (1998) Micropropagation of licorice (Glycyrrhiza glabra l.) through shoot tip and nodal cultures. In Vitro CeIl Dev Biol Plant 34:331–334
Toivonen L, Rosenqvist H (1995) Establishment and growth characteristics of Glycyrrhiza glabra hairy root cultures. Plant Cell Tiss Org Cult 41:249–258
Wongwicha W, Tanaka H, Shoyama Y, Tuvshintogtokh I, Putalun W (2008) Production of glycyrrhizin in callus cultures of licorice. Z Naturforsch 63:413–417
Yoo S, Kim S (1976) Studies in tissue culture of medicinal plants (II). Tissue culture of Glycyrrhiza glabra L. var. glandulifera Reg. et Herder. Kor J Pharmacol 7:55D57
Zhang MZ, Jie X, Bing Y, Huiyong Y, Qiuyin C, Martha J, Shrubsole XC, Valentina K, Wei Z, Ambra P, Harris RC (2009) Inhibition of 11β-hydroxysteroid dehydrogenase type II selectively blocks the tumor COX-2 pathway and suppresses colon carcinogenesis in mice and humans. J Clin Invest 119:876–885
Zietkiewicz E, Rafalski A, Labuda D (1994) Genome fingerprinting by simple sequence repeat (SSR)-anchored polymerase chain reaction amplification. Genomics 20:176–183
Acknowledgments
This work was carried out under Research Council approved project MLP-3012 (Work Package 4). Authors are grateful to Members Research Council, for their valuable comments during the course of the study. We thank Mr. Rajender Gupta for HPLC and Mr. Sunil Kumar for photography.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by S. Werbrouck.
Rights and permissions
About this article
Cite this article
Gupta, S., Pandotra, P., Gupta, A.P. et al. Direct rhizogenesis, in vitro stolon proliferation and high-throughput regeneration of plantlets in Glycyrrhiza glabra . Acta Physiol Plant 35, 2699–2705 (2013). https://doi.org/10.1007/s11738-013-1302-1
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11738-013-1302-1